Dopamina

Escrito y verificado por la psicóloga Paula Villasante
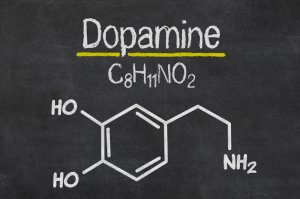
La dopamina es el neurotransmisor catecolaminérgico más importante del sistema nervioso central (SNC). Este neurotransmisor participa en la regulación de diversas funciones como la conducta motora, la emotividad y la afectividad, además de en la comunicación neuroendocrina (1).
La dopamina se sintetiza a partir del aminoácido L-tirosina. Además, existen mecanismos que regulan de manera muy precisa su síntensis y liberación.
Así, las técnicas de clonación molecular han permitido la identificación de 5 tipos de receptores dopaminérgicos, todos ellos acoplados a proteínas G y divididos en dos familias farmacológicas denominadas D1 y D2.
Sistemas dopaminérgicos
Las neuronas dopaminérgicas y sus proyecciones pueden agruparse en 3 sistemas principales (2):
Sistemas ultracortos
Los sistemas ultracortos son dos. El primero está formado por las células dopaminérgicas del bulbo olfatorio. Mientras, el segundo sistema lo componen las neuronas interflexiformes (similares a las amácrinas) presentes entre las capas plexiformes interna y externa de la retina.
Sistemas de longitud intermedia
Los sistemas de longitud intermedia incluyen:
a) El sistema tuberohipofisario, con origen en las células dopaminérgicas localizadas en los núcleos hipotalámicos arqueado y periventricular.
b) Neuronas localizadas en el hipotálamo dorsal y posterior. Estas envían proyecciones al hipotálamo dorsal anterior y a los núcleos septolaterales.
c) El grupo periventricular medular, que incluye a las neuronas dopaminérgicas localizadas en la periferia de los núcleos del tracto solitario y motor dorsal del nervio vago. También incluye a las células dispersas en la prolongación tegmental de la materia gris periacueductal.
Sistemas largos
Este grupo incluye a las neuronas de la región retrorubral, del área tegmental ventral y de la sustancia negra compacta. Estas envían proyecciones a tres regiones principales:
- El neoestriado (núcleos caudado y putamen).
- La corteza límbica (entorrinal, prefrontal medial y cíngulo).
- Otras estructuras límbicas (como el septum, el tubérculo olfatorio, el núcleo accumens, la amígdala y la corteza piriforme). Dentro de este grupo se encuentran además dos de las vías dopaminérgicas más importantes. Estas son la vía nigroestriatal y la vía mesolímbica.
Síntesis de la dopamina
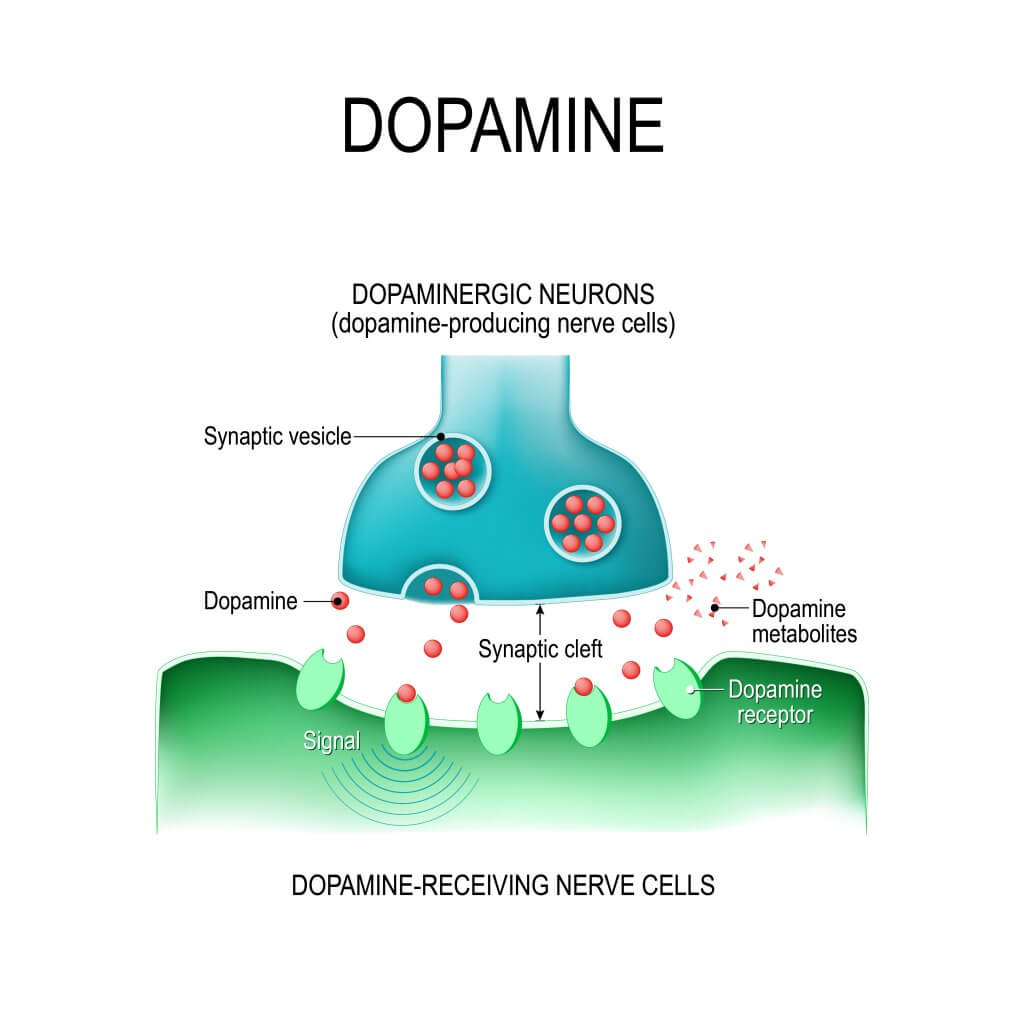
La síntesis del neurotransmisor tiene lugar en las terminales nerviosas dopaminérgicas donde se encuentran en alta concentración las enzimas responsables, la tirosina hidroxilasa (TH) y la descarboxilasa de aminoácidos aromáticos o L-DOPA descarboxilasa (2, 3).
Los trabajos de Nagatsu y col. (4) y de Levitt y col. (5) demostraron que la hidroxilación del aminoácido L-tirosina es el punto de regulación de la síntesis de catecolaminas en el SNC. Así, en consecuencia, la TH es la enzima limitante de la síntesis de la dopamina, la noradrenalina y la adrenalina.
La TH es un péptido de 498 aminoácidos presente de manera predominante en la fracción citosólica de las terminales catecolaminérgicas (6).
Liberación de la dopamina
En las terminales dopaminérgicas el neurotransmisor es sintetizado en el citoplasma de donde puede ser liberada directamente al espacio sináptico o bien ser transportada al interior de las vesículas sinápticas para ser liberada por exocitosis.
Liberación por exocitosis
En este proceso la dopamina contenida en vesículas es liberada al exterior al fusionarse la membrana vesicular con la membrana de la terminal presináptica. Este mecanismo está constituido por varias etapas.
En primer lugar, las vesículas transportan el neurotransmisor a su interior mediante una proteína transportadora con 12 dominios transmembranales que utiliza un gradiente electroquímico generado por una bomba (ATPasa) de protones (H+) (7).
Liberación independiente de Calcio
Este segundo tipo de liberación de dopamina es característicamente inhibido por fármacos que bloquean el transportador de dopamina presente en la membrana de la terminal sináptica.
La función de este transportador es terminar la acción del neurotransmisor, capturándolo hacia el interior de la terminal. Así, bajo ciertas condiciones el transportador opera en sentido inverso liberando dopamina al exterior (8).
Síntesis de dopamina y liberación
Se ha observado que la dopamina que se libera de manera preferente en respuesta a estimulación sináptica es la recién sintetizada.
El neurotransmisor parece así encontrarse en dos pozas metabólicas, ambas vesiculares; una que contiene a la dopamina recién sintetizada y una segunda que correspondería a una poza que funciona como almacén (9).
Regulación de la liberación de dopamina
La regulación de la liberación de dopamina puede producirse mediante dos tipos de regulaciones. Estas son:
- Regulación por autorreceptores.
- Regulación por heterorreceptores.
Receptores dopaminérgicos
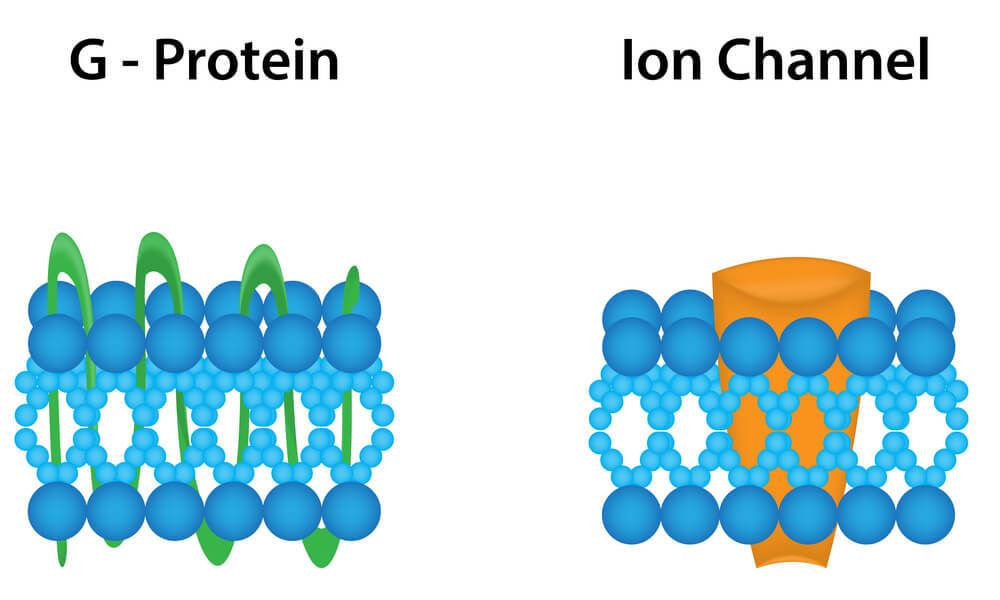
Actualmente, los receptores se definen como moléculas o composiciones moleculares que pueden reconocer selectivamente a un ligando (agonista o antagonista) y ser activados por el ligando con eficacia intrínseca (agonista) para iniciar un evento celular (10).
Los receptores para la dopamina pertenecen a la superfamilia de receptores (con más de 100 miembros) acoplados a proteínas G.
En esta familia de receptores, el reconocimiento del neurotransmisor y la molécula efectora (típicamente una enzima que produce un segundo mensajero difusible) son entidades diferentes, acopladas entre sí por una proteína con capacidad para unir nucleótidos de guanina (proteína G) (11).
La dopamina es un neurotransmisor de gran importancia en el sistema nervioso central. Los efectos funcionales de la dopamina se ejercen a través de la activación de 5 subtipos de receptores, todos ellos acoplados a proteínas G y agrupados en dos familias farmacológicas, D1 (D1 y D5) y D2 (D2, D3, D4).
Alteraciones diversas del SNC (enfermedad de Parkinson, esquizofrenia y adicción a drogas, entre otras) se han relacionado con transtornos de la transmisión dopaminérgica.
En consecuencia, el estudio de los aspectos aún no entendidos de la función de la dopamina, de los diferentes subtipos de receptores y de los mecanismos de transducción de señales, permitirá no sólo avanzar en la comprensión de la función de los sistemas dopaminérgicos, sino también diseñar nuevas estrategias farmacológicas que incidan en la terapéutica de dichos procesos patológicos (1).
La dopamina es el neurotransmisor catecolaminérgico más importante del sistema nervioso central (SNC). Este neurotransmisor participa en la regulación de diversas funciones como la conducta motora, la emotividad y la afectividad, además de en la comunicación neuroendocrina (1).
La dopamina se sintetiza a partir del aminoácido L-tirosina. Además, existen mecanismos que regulan de manera muy precisa su síntensis y liberación.
Así, las técnicas de clonación molecular han permitido la identificación de 5 tipos de receptores dopaminérgicos, todos ellos acoplados a proteínas G y divididos en dos familias farmacológicas denominadas D1 y D2.
Sistemas dopaminérgicos
Las neuronas dopaminérgicas y sus proyecciones pueden agruparse en 3 sistemas principales (2):
Sistemas ultracortos
Los sistemas ultracortos son dos. El primero está formado por las células dopaminérgicas del bulbo olfatorio. Mientras, el segundo sistema lo componen las neuronas interflexiformes (similares a las amácrinas) presentes entre las capas plexiformes interna y externa de la retina.
Sistemas de longitud intermedia
Los sistemas de longitud intermedia incluyen:
a) El sistema tuberohipofisario, con origen en las células dopaminérgicas localizadas en los núcleos hipotalámicos arqueado y periventricular.
b) Neuronas localizadas en el hipotálamo dorsal y posterior. Estas envían proyecciones al hipotálamo dorsal anterior y a los núcleos septolaterales.
c) El grupo periventricular medular, que incluye a las neuronas dopaminérgicas localizadas en la periferia de los núcleos del tracto solitario y motor dorsal del nervio vago. También incluye a las células dispersas en la prolongación tegmental de la materia gris periacueductal.
Sistemas largos
Este grupo incluye a las neuronas de la región retrorubral, del área tegmental ventral y de la sustancia negra compacta. Estas envían proyecciones a tres regiones principales:
- El neoestriado (núcleos caudado y putamen).
- La corteza límbica (entorrinal, prefrontal medial y cíngulo).
- Otras estructuras límbicas (como el septum, el tubérculo olfatorio, el núcleo accumens, la amígdala y la corteza piriforme). Dentro de este grupo se encuentran además dos de las vías dopaminérgicas más importantes. Estas son la vía nigroestriatal y la vía mesolímbica.
Síntesis de la dopamina
La síntesis del neurotransmisor tiene lugar en las terminales nerviosas dopaminérgicas donde se encuentran en alta concentración las enzimas responsables, la tirosina hidroxilasa (TH) y la descarboxilasa de aminoácidos aromáticos o L-DOPA descarboxilasa (2, 3).
Los trabajos de Nagatsu y col. (4) y de Levitt y col. (5) demostraron que la hidroxilación del aminoácido L-tirosina es el punto de regulación de la síntesis de catecolaminas en el SNC. Así, en consecuencia, la TH es la enzima limitante de la síntesis de la dopamina, la noradrenalina y la adrenalina.
La TH es un péptido de 498 aminoácidos presente de manera predominante en la fracción citosólica de las terminales catecolaminérgicas (6).
Liberación de la dopamina
En las terminales dopaminérgicas el neurotransmisor es sintetizado en el citoplasma de donde puede ser liberada directamente al espacio sináptico o bien ser transportada al interior de las vesículas sinápticas para ser liberada por exocitosis.
Liberación por exocitosis
En este proceso la dopamina contenida en vesículas es liberada al exterior al fusionarse la membrana vesicular con la membrana de la terminal presináptica. Este mecanismo está constituido por varias etapas.
En primer lugar, las vesículas transportan el neurotransmisor a su interior mediante una proteína transportadora con 12 dominios transmembranales que utiliza un gradiente electroquímico generado por una bomba (ATPasa) de protones (H+) (7).
Liberación independiente de Calcio
Este segundo tipo de liberación de dopamina es característicamente inhibido por fármacos que bloquean el transportador de dopamina presente en la membrana de la terminal sináptica.
La función de este transportador es terminar la acción del neurotransmisor, capturándolo hacia el interior de la terminal. Así, bajo ciertas condiciones el transportador opera en sentido inverso liberando dopamina al exterior (8).
Síntesis de dopamina y liberación
Se ha observado que la dopamina que se libera de manera preferente en respuesta a estimulación sináptica es la recién sintetizada.
El neurotransmisor parece así encontrarse en dos pozas metabólicas, ambas vesiculares; una que contiene a la dopamina recién sintetizada y una segunda que correspondería a una poza que funciona como almacén (9).
Regulación de la liberación de dopamina
La regulación de la liberación de dopamina puede producirse mediante dos tipos de regulaciones. Estas son:
- Regulación por autorreceptores.
- Regulación por heterorreceptores.
Receptores dopaminérgicos
Actualmente, los receptores se definen como moléculas o composiciones moleculares que pueden reconocer selectivamente a un ligando (agonista o antagonista) y ser activados por el ligando con eficacia intrínseca (agonista) para iniciar un evento celular (10).
Los receptores para la dopamina pertenecen a la superfamilia de receptores (con más de 100 miembros) acoplados a proteínas G.
En esta familia de receptores, el reconocimiento del neurotransmisor y la molécula efectora (típicamente una enzima que produce un segundo mensajero difusible) son entidades diferentes, acopladas entre sí por una proteína con capacidad para unir nucleótidos de guanina (proteína G) (11).
La dopamina es un neurotransmisor de gran importancia en el sistema nervioso central. Los efectos funcionales de la dopamina se ejercen a través de la activación de 5 subtipos de receptores, todos ellos acoplados a proteínas G y agrupados en dos familias farmacológicas, D1 (D1 y D5) y D2 (D2, D3, D4).
Alteraciones diversas del SNC (enfermedad de Parkinson, esquizofrenia y adicción a drogas, entre otras) se han relacionado con transtornos de la transmisión dopaminérgica.
En consecuencia, el estudio de los aspectos aún no entendidos de la función de la dopamina, de los diferentes subtipos de receptores y de los mecanismos de transducción de señales, permitirá no sólo avanzar en la comprensión de la función de los sistemas dopaminérgicos, sino también diseñar nuevas estrategias farmacológicas que incidan en la terapéutica de dichos procesos patológicos (1).
-
Bahena-Trujillo, R., Flores, G., & Arias-Montaño, J. A. (2000). Dopamina: síntesis, liberación y receptores en el Sistema Nervioso Central. Revista Biomédica, 11(1), 39-60.
-
Cooper, J. R., Bloom, F. E., & Roth, R. H. (2003). The biochemical basis of neuropharmacology. Oxford University Press, USA.
-
Freund, T. F., Powell, J. F., & Smith, A. D. (1984). Tyrosine hydroxylase-immunoreactive boutons in synaptic contact with identified striatonigral neurons, with particular reference to dendritic spines. Neuroscience, 13(4), 1189-1215.
-
Nagatsu, T., Levitt, M., & Udenfriend, S. (1964). Tyrosine hydroxylase the initial step in norepinephrine biosynthesis. Journal of Biological Chemistry, 239(9), 2910-2917.
-
Levitt, M., Spector, S., Sjoerdsma, A., & Udenfriend, S. (1965). Elucidation of the rate-limiting step in norepinephrine biosynthesis in the perfused guinea-pig heart. Journal of Pharmacology and Experimental Therapeutics, 148(1), 1-8.
- Weiner N, Molinoff PB. Cathecolamines. (1989) En: Siegel GJ, Agranoff B, Albers RW, Molinoff PB eds. Basic Neurochemistry. 4th Ed. New York: Raven Press;. p. 233-51.
- Receptor and ion channel nomenclature. Trends Pharmacol Sci 1998.
-
Raiteri, M., Marchi, M., & Maura, G. (1982). Presynaptic muscarinic receptors increase striatal dopamine release evoked by ‘quasi-physiological’depolarization. European journal of pharmacology, 83(1-2), 127-129.
-
Feldman, R. S., Meyer, J. S., Quenzer, L. F., & Cooper, J. R. (1997). Principles of neuropsychopharmacology (No. 615+ 616 FEL). Sunderland: Sinauer Associates.
-
Humphrey, P. P. (1997). The characterization and classification of neurotransmitter receptors. Annals of the New York Academy of Sciences, 812(1), 1-13.
-
Kupfermann, I., Kandel, E. R., Schwartz, J. H., & Jessel, T. M. (1991). Principles of neural science.
Este texto se ofrece únicamente con propósitos informativos y no reemplaza la consulta con un profesional. Ante dudas, consulta a tu especialista.