Dopamine: systèmes dopaminergiques, synthèse et récepteurs

Escrito y verificado por la psicóloga Paula Villasante
La dopamine est le neurotransmetteur catécholaminergique le plus important du système nerveux central (SNC). Ce neurotransmetteur participe à la régulation de diverses fonctions telles que le comportement moteur, l’émotion et l’affectivité, ainsi qu’à la communication neuroendocrinienne (1).
Par ailleurs, la dopamine est synthétisée à partir de l’acide aminé L-tyrosine. De plus, il existe des mécanismes qui régulent très précisément sa synthèse et sa libération.
Ainsi, les techniques de clonage moléculaire ont permis d’identifier 5 types de récepteurs dopaminergiques, tous couplés à des protéines G et répartis en deux familles pharmacologiques appelées D1 et D2.
Systèmes dopaminergiques
Les neurones dopaminergiques et leurs projections peuvent être regroupés en 3 systèmes principaux (2):
Systèmes ultracourts
Il existe deux systèmes ultracourts. Le premier se forme par les cellules dopaminergiques du bulbe olfactif. Pendant ce temps, le deuxième système se compose de neurones interflexiformes (similaires aux amacrines) présents entre les couches plexiformes interne et externe de la rétine.
Systèmes de longueur intermédiaire
Les systèmes de longueur intermédiaire comprennent:
a) Le système tubéro-hypophysaire, provenant des cellules dopaminergiques situées dans les noyaux hypothalamiques arqués et périventriculaires.
b) Neurones situés dans l’hypothalamus dorsal et postérieur. Ceux-ci envoient des projections vers l’hypothalamus dorsal antérieur et vers les noyaux septaux-latéraux.
c) Le groupe périventriculaire médullaire, qui comprend les neurones dopaminergiques situés à la périphérie des noyaux du tractus moteur solitaire et dorsal du nerf vague. Il comprend également des cellules dispersées dans le prolongement tegmental de la matière grise périaqueducale.
Systèmes longs
Ce groupe comprend les neurones de l’aire tegmentale ventrale et de la substance noire compact. Ils envoient des projections à trois régions principales:
- Le néostriatum (noyaux caudés et putamen).
- Le cortex limbique (entorhinal, préfrontal médial et cingulum).
- Autres structures limbiques (telles que le septum, le tubercule olfactif, le noyau accumbens, l’amygdale et le cortex piriforme). Au sein de ce groupe, il existe également deux des voies dopaminergiques les plus importantes. Ce sont la voie nigrostriatale et la voie mésolimbique.
Synthèse de la dopamine
La synthèse du neurotransmetteur a lieu dans les terminaisons nerveuses dopaminergiques où les enzymes responsables, la tyrosine hydroxylase (TH) et la décarboxylase des acides aminés aromatiques ou L-DOPA décarboxylase se trouvent en forte concentration (2, 3).
Les travaux de Nagatsu et al. (4) et Levitt et al. (5) ont démontré que l’hydroxylation de l’acide aminé L-tyrosine est le point de régulation de la synthèse des catécholamines dans le SNC. Par conséquent, la TH est l’enzyme limitante pour la synthèse de la dopamine, de la norépinéphrine et de l’épinéphrine.
TH est un peptide de 498 acides aminés présent majoritairement dans la fraction cytosolique des terminaisons catécholaminergiques (6).
Libération de dopamine
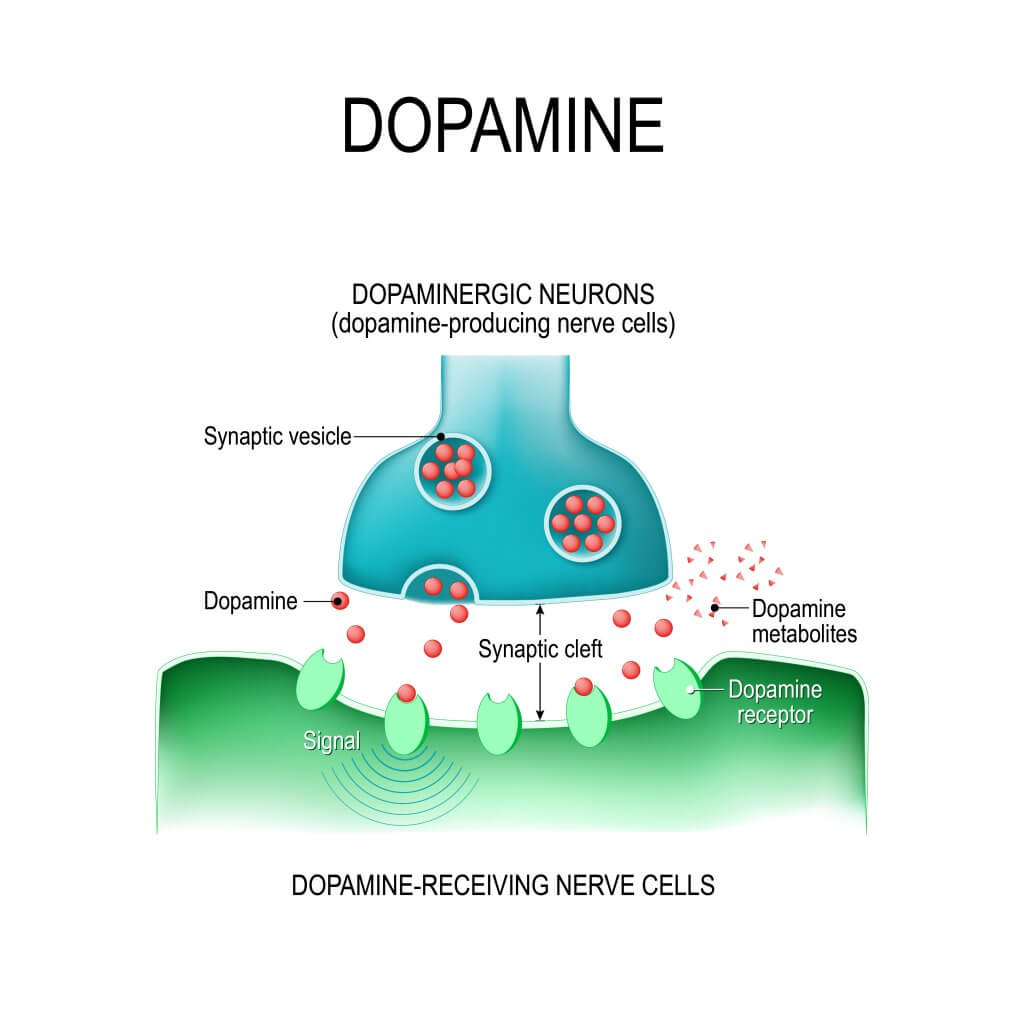
Aux terminaisons dopaminergiques, le neurotransmetteur est synthétisé dans le cytoplasme d’où il peut être libéré directement dans l’espace synaptique ou être transporté dans les vésicules synaptiques pour être libéré par exocytose.
Libération de la dopamine par exocytose
Dans ce processus, la dopamine contenue dans les vésicules est libérée vers l’extérieur lorsque la membrane vésiculaire fusionne avec la membrane de la terminaison présynaptique. Ce mécanisme se compose de plusieurs étapes.
Premièrement, les vésicules transportent le neurotransmetteur à l’intérieur à travers une protéine de transport avec 12 domaines transmembranaires qui utilisent un gradient électrochimique généré par une pompe à protons (H +) (ATPase) (7).
Libération indépendante de calcium
Ce deuxième type de libération de dopamine est inhibé de manière caractéristique par des médicaments qui bloquent le transporteur de dopamine présent dans la membrane terminale synaptique.
La fonction de ce transporteur est de mettre fin à l’action du neurotransmetteur, en le capturant dans le terminal. Ainsi, dans certaines conditions, le transporteur fonctionne en sens inverse, libérant de la dopamine vers l’extérieur (8).
Synthèse et libération de dopamine
La dopamine qui est principalement libérée en réponse à une stimulation synaptique s’est avérée être nouvellement synthétisée.
Le neurotransmetteur semble donc se trouver dans deux pools métaboliques, tous deux vésiculaires. L’un qui contient de la dopamine nouvellement synthétisée et un second qui correspondrait à un pool qui fonctionne comme un magasin (9).
Régulation de la libération de dopamine
La régulation de la libération de dopamine peut se faire par deux types de régulation. Ceux-ci sont:
- Régulation par autorécepteurs.
- Régulation par les hétérorécepteurs.
Récepteurs de la dopamine
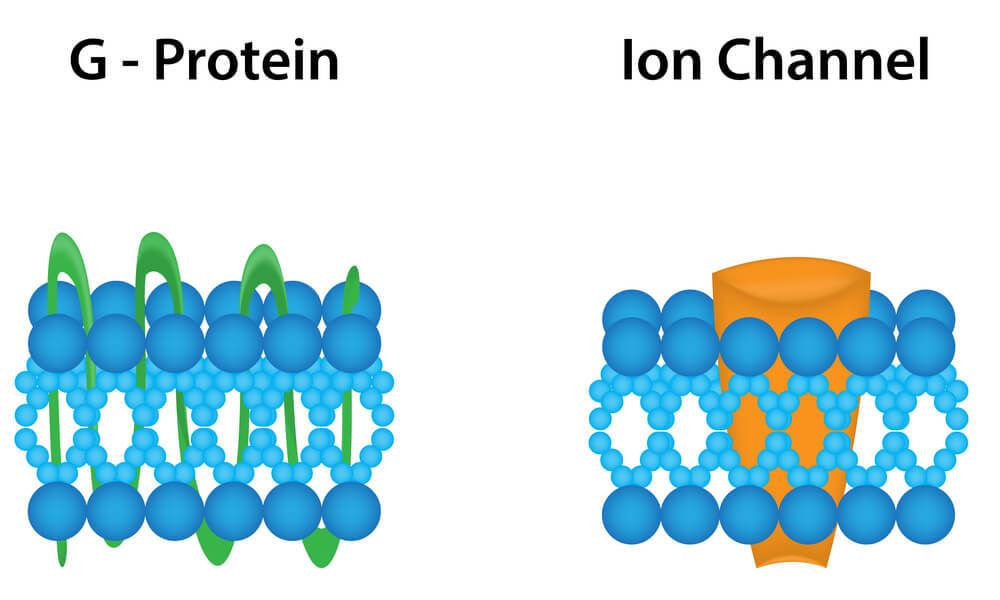
Actuellement, les récepteurs sont définis comme des molécules ou des compositions moléculaires qui peuvent reconnaître sélectivement un ligand (agoniste ou antagoniste) et être activés par le ligand avec une efficacité intrinsèque (agoniste) pour initier un événement cellulaire (10).
Les récepteurs de la dopamine appartiennent à la superfamille des récepteurs (avec plus de 100 membres) couplés aux protéines G.
Dans cette famille de récepteurs, la reconnaissance du neurotransmetteur et la molécule effectrice (typiquement une enzyme qui produit un second messager diffusible) sont des entités différentes, couplées ensemble par une protéine ayant la capacité de lier les nucléotides guanines (protéine G) (11).
La dopamine est un neurotransmetteur très important dans le système nerveux central. Les effets fonctionnels de la dopamine s’exercent par l’activation de 5 sous-types de récepteurs, tous couplés à des protéines G et regroupés en deux familles pharmacologiques, D1 (D1 et D5) et D2 (D2, D3, D4).
Diverses altérations du SNC (maladie de Parkinson, schizophrénie et toxicomanie, entre autres) ont été liées à des troubles de la transmission de la dopamine.
Par conséquent, l’étude d’aspects non encore compris de la fonction dopaminergique, des différents sous-types de récepteurs et des mécanismes de transduction du signal, permettra non seulement d’avancer dans la compréhension de la fonction des systèmes dopaminergiques, mais également de concevoir de nouvelles stratégies pharmacologiques qui affectent le traitement de ces processus pathologiques (1).
La dopamine est le neurotransmetteur catécholaminergique le plus important du système nerveux central (SNC). Ce neurotransmetteur participe à la régulation de diverses fonctions telles que le comportement moteur, l’émotion et l’affectivité, ainsi qu’à la communication neuroendocrinienne (1).
Par ailleurs, la dopamine est synthétisée à partir de l’acide aminé L-tyrosine. De plus, il existe des mécanismes qui régulent très précisément sa synthèse et sa libération.
Ainsi, les techniques de clonage moléculaire ont permis d’identifier 5 types de récepteurs dopaminergiques, tous couplés à des protéines G et répartis en deux familles pharmacologiques appelées D1 et D2.
Systèmes dopaminergiques
Les neurones dopaminergiques et leurs projections peuvent être regroupés en 3 systèmes principaux (2):
Systèmes ultracourts
Il existe deux systèmes ultracourts. Le premier se forme par les cellules dopaminergiques du bulbe olfactif. Pendant ce temps, le deuxième système se compose de neurones interflexiformes (similaires aux amacrines) présents entre les couches plexiformes interne et externe de la rétine.
Systèmes de longueur intermédiaire
Les systèmes de longueur intermédiaire comprennent:
a) Le système tubéro-hypophysaire, provenant des cellules dopaminergiques situées dans les noyaux hypothalamiques arqués et périventriculaires.
b) Neurones situés dans l’hypothalamus dorsal et postérieur. Ceux-ci envoient des projections vers l’hypothalamus dorsal antérieur et vers les noyaux septaux-latéraux.
c) Le groupe périventriculaire médullaire, qui comprend les neurones dopaminergiques situés à la périphérie des noyaux du tractus moteur solitaire et dorsal du nerf vague. Il comprend également des cellules dispersées dans le prolongement tegmental de la matière grise périaqueducale.
Systèmes longs
Ce groupe comprend les neurones de l’aire tegmentale ventrale et de la substance noire compact. Ils envoient des projections à trois régions principales:
- Le néostriatum (noyaux caudés et putamen).
- Le cortex limbique (entorhinal, préfrontal médial et cingulum).
- Autres structures limbiques (telles que le septum, le tubercule olfactif, le noyau accumbens, l’amygdale et le cortex piriforme). Au sein de ce groupe, il existe également deux des voies dopaminergiques les plus importantes. Ce sont la voie nigrostriatale et la voie mésolimbique.
Synthèse de la dopamine
La synthèse du neurotransmetteur a lieu dans les terminaisons nerveuses dopaminergiques où les enzymes responsables, la tyrosine hydroxylase (TH) et la décarboxylase des acides aminés aromatiques ou L-DOPA décarboxylase se trouvent en forte concentration (2, 3).
Les travaux de Nagatsu et al. (4) et Levitt et al. (5) ont démontré que l’hydroxylation de l’acide aminé L-tyrosine est le point de régulation de la synthèse des catécholamines dans le SNC. Par conséquent, la TH est l’enzyme limitante pour la synthèse de la dopamine, de la norépinéphrine et de l’épinéphrine.
TH est un peptide de 498 acides aminés présent majoritairement dans la fraction cytosolique des terminaisons catécholaminergiques (6).
Libération de dopamine
Aux terminaisons dopaminergiques, le neurotransmetteur est synthétisé dans le cytoplasme d’où il peut être libéré directement dans l’espace synaptique ou être transporté dans les vésicules synaptiques pour être libéré par exocytose.
Libération de la dopamine par exocytose
Dans ce processus, la dopamine contenue dans les vésicules est libérée vers l’extérieur lorsque la membrane vésiculaire fusionne avec la membrane de la terminaison présynaptique. Ce mécanisme se compose de plusieurs étapes.
Premièrement, les vésicules transportent le neurotransmetteur à l’intérieur à travers une protéine de transport avec 12 domaines transmembranaires qui utilisent un gradient électrochimique généré par une pompe à protons (H +) (ATPase) (7).
Libération indépendante de calcium
Ce deuxième type de libération de dopamine est inhibé de manière caractéristique par des médicaments qui bloquent le transporteur de dopamine présent dans la membrane terminale synaptique.
La fonction de ce transporteur est de mettre fin à l’action du neurotransmetteur, en le capturant dans le terminal. Ainsi, dans certaines conditions, le transporteur fonctionne en sens inverse, libérant de la dopamine vers l’extérieur (8).
Synthèse et libération de dopamine
La dopamine qui est principalement libérée en réponse à une stimulation synaptique s’est avérée être nouvellement synthétisée.
Le neurotransmetteur semble donc se trouver dans deux pools métaboliques, tous deux vésiculaires. L’un qui contient de la dopamine nouvellement synthétisée et un second qui correspondrait à un pool qui fonctionne comme un magasin (9).
Régulation de la libération de dopamine
La régulation de la libération de dopamine peut se faire par deux types de régulation. Ceux-ci sont:
- Régulation par autorécepteurs.
- Régulation par les hétérorécepteurs.
Récepteurs de la dopamine
Actuellement, les récepteurs sont définis comme des molécules ou des compositions moléculaires qui peuvent reconnaître sélectivement un ligand (agoniste ou antagoniste) et être activés par le ligand avec une efficacité intrinsèque (agoniste) pour initier un événement cellulaire (10).
Les récepteurs de la dopamine appartiennent à la superfamille des récepteurs (avec plus de 100 membres) couplés aux protéines G.
Dans cette famille de récepteurs, la reconnaissance du neurotransmetteur et la molécule effectrice (typiquement une enzyme qui produit un second messager diffusible) sont des entités différentes, couplées ensemble par une protéine ayant la capacité de lier les nucléotides guanines (protéine G) (11).
La dopamine est un neurotransmetteur très important dans le système nerveux central. Les effets fonctionnels de la dopamine s’exercent par l’activation de 5 sous-types de récepteurs, tous couplés à des protéines G et regroupés en deux familles pharmacologiques, D1 (D1 et D5) et D2 (D2, D3, D4).
Diverses altérations du SNC (maladie de Parkinson, schizophrénie et toxicomanie, entre autres) ont été liées à des troubles de la transmission de la dopamine.
Par conséquent, l’étude d’aspects non encore compris de la fonction dopaminergique, des différents sous-types de récepteurs et des mécanismes de transduction du signal, permettra non seulement d’avancer dans la compréhension de la fonction des systèmes dopaminergiques, mais également de concevoir de nouvelles stratégies pharmacologiques qui affectent le traitement de ces processus pathologiques (1).
-
Bahena-Trujillo, R., Flores, G., & Arias-Montaño, J. A. (2000). Dopamina: síntesis, liberación y receptores en el Sistema Nervioso Central. Revista Biomédica, 11(1), 39-60.
-
Cooper, J. R., Bloom, F. E., & Roth, R. H. (2003). The biochemical basis of neuropharmacology. Oxford University Press, USA.
-
Freund, T. F., Powell, J. F., & Smith, A. D. (1984). Tyrosine hydroxylase-immunoreactive boutons in synaptic contact with identified striatonigral neurons, with particular reference to dendritic spines. Neuroscience, 13(4), 1189-1215.
-
Nagatsu, T., Levitt, M., & Udenfriend, S. (1964). Tyrosine hydroxylase the initial step in norepinephrine biosynthesis. Journal of Biological Chemistry, 239(9), 2910-2917.
-
Levitt, M., Spector, S., Sjoerdsma, A., & Udenfriend, S. (1965). Elucidation of the rate-limiting step in norepinephrine biosynthesis in the perfused guinea-pig heart. Journal of Pharmacology and Experimental Therapeutics, 148(1), 1-8.
- Weiner N, Molinoff PB. Cathecolamines. (1989) En: Siegel GJ, Agranoff B, Albers RW, Molinoff PB eds. Basic Neurochemistry. 4th Ed. New York: Raven Press;. p. 233-51.
- Receptor and ion channel nomenclature. Trends Pharmacol Sci 1998.
-
Raiteri, M., Marchi, M., & Maura, G. (1982). Presynaptic muscarinic receptors increase striatal dopamine release evoked by ‘quasi-physiological’depolarization. European journal of pharmacology, 83(1-2), 127-129.
-
Feldman, R. S., Meyer, J. S., Quenzer, L. F., & Cooper, J. R. (1997). Principles of neuropsychopharmacology (No. 615+ 616 FEL). Sunderland: Sinauer Associates.
-
Humphrey, P. P. (1997). The characterization and classification of neurotransmitter receptors. Annals of the New York Academy of Sciences, 812(1), 1-13.
-
Kupfermann, I., Kandel, E. R., Schwartz, J. H., & Jessel, T. M. (1991). Principles of neural science.
Este texto se ofrece únicamente con propósitos informativos y no reemplaza la consulta con un profesional. Ante dudas, consulta a tu especialista.